| томский государственный университет |
| кафедра органической химии |
|
экспрессия векторов в эукариотических
клетках
|
Многие генетические заболевания
у млекопитающих связаны с отсутствием или дефектами внутриклеточных белков,
вызванные дефектами генов, кодирующих эти белки. Лечение таких заболеваний возможно
с использованием рекомбинантных технологий, с применением специально сконструированных
кольцевых ДНК (векторов), содержащих НОРМАЛЬНЫЕ гены, отсутствующие в организме.
К настоящему моменту времени созданы многочисленные векторы для этих целей,
способные экспрессировать в клетках млекопитающих in vitro. Однако,
подобные векторы, применяемые in vivo,
должны быть селективны по отношению к тканям с дефектными клетками. Непосредственное
генетическое модифицирование в организме насчитывает слишком мало примеров успешного
применения.
Определенные типы разработанных
векторов позволяют репликацию, транскрипцию и трансляцию чужеродных генов, осуществляемую
на клеточных культурах, выращиваемых in vitro,
включая ДНК- и РНК- вирусные векторы, содержащие вставки целевой ДНК. Эти вирусные
векторы способны инфицировать и реплицировать в клетке-хозяине. Экспериментально
сконструированные векторы позволяют экспрессию чужеродных генов только, если
содержат существенные элементы ДНК. Обычно эти элементы привносятся непосредственно
с вирусной ДНК.
Так называемые челночные
векторы, содержат сигналы репликации,
необходимые в бактериальных системах и системах млекопитающих, поэтому могут
применяться для производства белков в обоих типах клеток. Челночные векторы
могут быть клонированы и очищены из бактериальных клеток в больших количествах,
а затем перенесены для экспрессии в клетки млекопитающих. Некоторые из них способны
внедряться в геном клетки-хозяина, другие- остаются внехромосомальными
(эписомальными), сохраняя, однако
стабильную возможность экспрессии заложенного в них интересуемого гена, Некоторые
из последних- кочуя в дочерние клетки при их делении, другие- работая только
в период жизни клетки.
Процесс переноса информации чужеродной ДНК в эукариоту в виде вектора называется трансфекцией. Подобный процесс переноса информации в клетки прокариот называют трансформацией. Наиболее часто используемый метод трансфекции включает образование комплекса ДНК с фосфатом кальция или DEAE-декстраном, которые затем захватываются клеткой в результате эндоцитоза. ДНК затем переносится из цитоплазмы в ядро, где и происходит ее репликация и экспрессия. Детали механизма трансфекции неизвестны. Обычно трансфекции подвергаются 10-20 % клеток культуры.
Экспрессия рекомбинантных генов в клетках млекопитающих требует
присутствия в векторе ДНК-контролирующих элементов, которые необязательны в
бактериальных системах. Для экспрессии в эукариотической клетке, ген должен
быть вставлен в вектор в нужном направлении относительно контролирующих элементов,
включающих промотер, сигналы полиаденилирования, в некоторых случаях- интроны.
Некоторые или все из этих элементов могут оказаться в векторе, если использовалась
цельная геномная ДНК при клонировании. Если же клонировали только часть ДНК,
они могут отсутствовать, поскольку были удалены при обработке рестриктазами.
В таких случаях они должны присутствовать в самом векторе.
Популяция эукариотических клеток, подвергшихся трансфекции, должна быть обнаруживаема с целью их направленного выращивания, ведь их относительное число не превышает 10-20 %. Конструирование векторов, содержащих экспрессируемый ген и маркер одновременно, чрезвычайно сложно, поэтому прибегают к котрансфекции, когда в клетку вносят одновременно два вектора, с геном и маркером. В большинстве случаев, не менее 90 % преобразованных клеток содержат оба вектора.
Два из наиболее широко применяемых
маркера- гены тимидинкиназы (тк)
и дигидрофолат-редуктазы (дгфр).
Продукт тк-гена
– тимидинкиназа образуется в большинстве клеток
млекопитающих и участвует в путях метаболизма тимидина. Некоторые мутантные
линии клеток не содержат функционального гена тимидинкиназы (тк-)
и такие клетки не выживают в средах, содержащих гипоксантин, аминоптерин и тимидин.
Только те из (тк-)-мутантных
клеток, которые в результате котрансфекции получат вектор, несущий нормальный
ген, способны к росту. Они и подвергаются процедуре внедрения интересуемого
генома.
Ген дегидрофолат-редуктазы (дгфр)
необходим для поддержания в клетке нормального
уровня активности дегидрофолат-редуктазы, необходимой в биосинтезе нуклеотидов.
Клетки, не содержащие этого фермента, выживают только в средах, содержащих тимидин,
глицин и пуриновые азотистые основания. Если
провести трансфекцию мутантных клеток одновременно двумя векторами (с интересуемым
геном и геном дгфр),
то становится возможным их определение от
нетрансфектированных клеток при выращивании на средах, не содержащих перечисленные
выше компоненты.
Другой подход для определения немутировавших клеток- использование для котрансфекции бактериального гена, кодирующего аминогликозид-3’-фосфотрансферазу (АФТ). Клетки, экспрессирующие ген АФТ, устойчивы к аминогликозидным антибиотикам типа неомицина и канамицина, ингибирующих синтез протеинов как у прокариот, так и у эукариот. Поэтому векторы, несущие ген АФТ, могут применяться как на бактериальных клетках, так и на клетках млекопитающих.
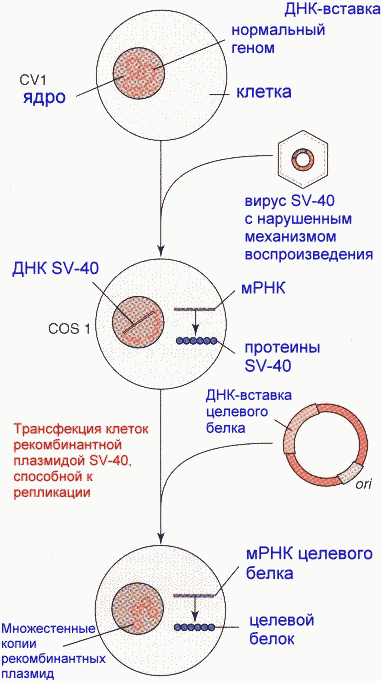 |
“Чужие” гены можно экспрессировать
применяя трансформированные вирусом эукариотические клетки. Например,
клетки линии COS-1 есть симиановые
клетки CV-1, трансформированные ori-дефектным
вирусом SV-40. Дефектный вирусный
геном внедряется в геном клетки и способен только продуцировать вирусные
белки, в то время как сами вирусы не продуцируются, вследствие нарушенной
репликативной функции. Однако, если в трансформированную таким образом
клетку внести рекомбинантную ДНК вируса SV-40
с ненарушенной репликативной системой, содержащую к тому же вставку из
интересуемой ДНК, то начнется репликация рекомбинантных вирусных плазмид
в больших количествах. Они, оставаясь внехромосомальными, накапливаются
в количествах до 100 тысяч на клетку, одновременно происходит транскрипция
интересуемой ДНК-овой вставки с образованием мРНК и синтез целевого белка
в результате трансляции. Клетки COS-1
погибают через 3-4 дня, возможно, из-за токсического действия эписомальной
векторной ДНК, и целевой белок может быть выделен.
|
вверх
процессинг нуклеиновых кислот
секвенирование ДНК
рекомбинантные ДНК и биотехнология
нуклеиновые кислоты
вернуться на начальную
страницу ORGCHEM.TSU.RU
подробное содержание раздела
|
© khassanov, MMII-MMXVIII
|