

При полном гидролизе нуклеиновых кислот (НК) выделены соединения:
а) пуриновые и пиримидиновые основания;
б) моносахариды РИБОЗА или ДЕЗОКСИРИБОЗА (С5Н10О5);
в) фосфорная кислота.
Строение моносахаридов:

Строение пуриновых оснований:

Строение пиримидиновых оснований:

Нуклеиновые кислоты имеют различающийся состав. В частности, дезоксирибонуклеиновые кислоты (ДНК) содержат дезоксирибозу, а рибонуклеиновые кислоты (РНК)- рибозу. Эти и другие отличия в составе нуклеиновых кислот приведены в таблице:
|
Одинаковые компоненты |
Отличающиеся компоненты |
|
|
ДНК |
РНК |
|
|
АДЕНИН ГУАНИН ЦИТОЗИН |
ДЕЗОКСИРИБОЗА ТИМИН |
РИБОЗА УРАЦИЛ |
НУКЛЕОЗИДЫ- соединения азотистого основания и углеводов (рибозы и дезоксирибозы). Нуклеозиды образуются за счет N-гликозидной связи между ДЕВЯТЫМ атомом азота у ПУРИНОВЫХ (первым атомом азота- у ПИРИМИДИНОВЫХ) оснований и гидроксилом ПЕРВОГО атома углерода рибозы или дезоксирибозы. Во избежание путаницы, нумерация атомов азотистых оснований осуществляется арабскими цифрами, а у атомов углерода рибоз- арабскими цифрами со “штрихом”.
| пуриновые | |
| пиримидиновые | |
НУКЛЕОТИДЫ отличаются от нуклеозидов наличием остатков фосфорной кислоты (от одного до трех), связанных простой эфирной связью с гидроксилом 5’ атома углерода рибоз. Остатки фосфорных кислот между собой также связаны простой эфирной связью. В зависимости от числа остатков фосфорной кислоты в нуклеотидах различают МОНО-, ДИ- и ТРИФОСФОНУКЛЕОТИДЫ. Их номенклатура приведена в таблице:
|
азотистые основания |
нуклеозиды |
Нуклеотиды |
|
|
полное название |
Сокращенное |
||
|
аденин |
аденозин |
Аденозинмонофосфат |
АМФ |
|
гуанин |
гуанозин |
Гуанозинмонофосфат |
ГМФ |
|
цитозин |
цитидин |
Цитидинмонофосфат |
ЦМФ |
|
урацил |
уридин |
Уридинмонофосфат |
УМФ |
|
тимин |
тимидин |
тимидинмонофосфат |
ТМФ |
 |
Собственно НУКЛЕИНОВЫЕ кислоты представляют собой полинуклеотидмонофосфаты. Полимерная цепь образуется за счет фосфодиэфирной связи между 3’- гидроксилом одного нуклеотида и 5’- гидроксилом другого. Таким образом, первичная структура нуклеиновых кислот представляет собой ПОРЯДОК чередования нуклеотидов в полинуклеотидной цепи. Один из концов этой цепи (изображаемый слева) имеет свободный гидроксил при 5’- атоме С, а другой (изображаемый справа)- свободный гидроксил при 3’- атоме углерода рибоз. Поскольку основой нуклеиновых кислот является САХАРОФОСФАТНЫЙ ОСТОВ, в сокращенных написаниях участков цепи используют лишь ОДНОБУКВЕННЫЕ СИМВОЛЫ соответствующего азотистого основания. Полное и схематичное обозначение участка полинуклеотидной цепи приведены ниже: 5’-НО-G-A-A-T-C-T-A-C-A-…3' Вследствие наличия сильнодиссоциирующих фосфатных групп, нуклеиновые кислоты (НК) легко образуют связи с оснОвными белками (высокое значение pI) с образованием НУКЛЕОПРОТЕИНОВ. Протеины отделяются от НК детергентами или после расщепления белков протеиназами НК осаждаются спиртом. |
Подобно белкам, ДНК имеют первичную, вторичную и третичную структуру.
ПЕРВИЧНАЯ СТРУКТУРА есть последовательность чередования нуклеотидов в цепи ДНК и РНК. Сложность расшифровки структуры связана с наличием всего 4-х видов нуклеотидов при их огромном числе в молекуле.
ПРАВИЛА ЧАРГАФФА
1) сумма ПУРИНОВЫХ оснований в молекуле ДНК всегда равна сумме ПИРИМИДИНОВЫХ оснований.
2) А=Т, А/Т= 1; 3) Г=Ц, Г/Ц=1; 4) А+Т=Г+Ц; 5) если А+Т>Г+Ц, то АТ-тип.
ВТОРИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Расшифрована впервые в 1953 г. Уотсоном и Крик (Watson, Crick).
Вторичная структура ДНК представляет собой свернутые в спираль ДВЕ комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи. Образование вторичной структуры НК возможно вследствие проявления эффектов КОМПЛЕМЕНТАРНОСТИ и СТЭКИНГ-ВЗАИМОДЕЙСТВИЙ. Очень часто наблюдаются двунитевые спирализованные молекулы ДНК, замкнутые в кольцо с ковалентно связанными концами. Они не имеют разрывов у каждой в отдельности полинуклеотидной цепи. Подобные кольцевые ДНК, как правило, СУПЕРСПИРАЛИЗОВАНЫ, то есть кольцо дополнительно закручено в спираль. Суперспирализация- ПРАВИЛО, а не ИСКЛЮЧЕНИЕ, при условии отсутствия разрывов в фосфодиэфирных связях полинуклеотидной цепи.
КОМПЛЕМЕНТАРНОСТЬ - последовательность нуклеотидов в одной цепи автоматически определяет строго соответствующую ей последовательность нуклеотидов в
КОМПЛЕМЕНТАРНОЙ ей цепи. Так, азотистое основание Аденин (А) всегда взаимодействует только с комплементарным ему азотистым основанием Тимин (Т) в молекулах ДНК.
Одновременно азотистые основания Гуанин (Г) одной цепи взаимодействует только с комплементарними им азотистыми основаниями Цитозин (Ц) другой цепи (как в ДНК, так и в РНК).
Комплементарность оснований обеспечивается системой водородных связей. В молекулах РНК, имеющих, в основном, однонитевую структуру, на отдельных участках, азотистые основания А взаимодействуют с
комплементарными им азотистыми основаниями Урацил (У).

Аналогично происходит взаимодействие в процессах транскрипции, когда на матрице ДНК синтезируется молекула РНК (матричная, транспортная и т.д.), и наоборот, когда при участии РЕВЕРС-ТРАНСКРИПТАЗЫ
происходит синтез кДНК на матрице РНК.
СТЭКИНГ-ВЗАИМОДЕЙСТВИЯ- особого рода (Ван-дер-Ваальсовы) взаимодействия между выложенными в стопку (как монеты) друг над другом азотистых оснований.
Имеются А, В, С и Z-формы двунитевых участков ДНК, отличающиеся наклонами плоскостей азотистых оснований друг относительно друга ( у А- 20о, В- 0о, С- 5о, Z- особая ломанная форма).

SBS (side-by-side)- форма (бок о бок). Две цепи расположены прямо и незакручены.
ТРЕТИЧНАЯ СТРУКТУРА ДНК
ДНК имеют формы ЛИНЕЙНАЯ, КОЛЬЦЕВАЯ, 2-х и 1-ЦЕПОЧЕЧНАЯ.
Двуцепочечные ДНК с "липкими" концами могут образовывать кольцо, которое далее ковалентно сшивается по сахарофосфатной цепи при помощи ДНК-лигазы.

Третичная структура ДНК у эукариотических клеток отличается тем, что многократная спирализация ДНК сопровождается образованием комплексов с белками.
46 хромосом (хроматид) человека
организованы в 23 пары. Средняя длина хромосомы составляет 130 млн. пар оснований
и имеет длину 5 см. Хромосома №1- 263 млн. п.о., хромосома № 46 –меньше 50 млн.
п.о. Если проложить все ДНК в В-конформации в линию, то их общая длина превысит
2 метра. Человеческая хромосома 16 имеет 2,5 мкм в длину, а длина самой ДНК-
3,7 см.
Понятно, что уместить такой длины
ДНК в ядре возможно только путем ее определенной упаковки. При образовании третичной
структуры ДНК человека происходит в среднем уменьшение ее размеров в 100 тысяч
раз.
 |
При образовании третичной структуры нуклеиновых кислот возможно образование ТРЕХ- и ЧЕТЫРЕХНИТЕВЫХ участков. Образование ТРЕХНИТЕВЫХ участков благодаря так называемым Хугстэновским взаимодействиям (Hoogsteen base pairs), когда одновременно взаимодействуют три основания: А-А-Т, Т-А-Т, Г-Г-Ц, Ц-Г-Ц. Аналогичным образом происходит образование ТЕТРАМЕРНЫХ участков ДНК. Точный биологический смысл появления трех- и четырехнитевых участков ДНК пока не выяснен. Имеются только лишь предположения о том, что такие участки возникают в местах, наиболее ответственных за процессы репликации и транскрипции. |
 |
Материал хромосом- ХРОМАТИН- содержит кроме самой ДНК также гистоны, негистоновые белки, небольшое количество РНК. Нуклеосомный кор содержит октамер гистонов (2 х (Н2а+Н2b+H3+H4)). ГИСТОН- простой белок (примерно 50% хроматина). Нуклеосомный кор образуется при оборачивании октамера гистонов двунитевой спирализованной ДНК на 1,5 оборота, отдельно включается дополнительный белок- гистон Н1. Все вместе называется ХРОМАТОСОМом .Н1- очень богат ЛИЗ, Н2а, Н2b- умеренное количество ЛИЗ, Н3- есть ЦИС, умеренно- АРГ, Н4- богат АРГ и ГЛИ. |
Хроматосомы образуются
на двунитевой спирали ДНК на дистанциях (называемых ЛИНКЕРами) от 20 до 90 пар
нуклеотидов и напоминают нанизанные на нитку бусины. Следующий этап- сворачивание
в спираль очень длинной последовательности “бус”. Эта спираль, в свою очередь,
претерпевает сворачивание в двужильные канаты, из которых образуются гроздья,
являющиеся небольшой частью хромосомы:

МАКРОМОЛЕКУЛЯРНАЯ СТРУКТУРА РНК, тРНК
РНК состоит из одной полинуклеотидной цепи, закрученной на себя, образует короткие двуспиральные шпильки в палиндромных участках (Г с Ц, А с У). тРНК- самые мелкие молекулы (ММ=23-30 тыс.) тРНК- переносчик аминокислот. Каждая тРНК переносит только одну аминокислоту, но на одну аминокислоту имеется более одной тРНК. Всего известно 61 тРНК.

Строение тРНК. Имеет АКЦЕПТИРУЮЩИЙ УЧАСТОК (присоединяет АК, при участии АТФ), ОБЩИЙ УЧАСТОК (петля дигидроуридина) обеспечивает связь со специфическим ферментом, ХАРАКТЕРНЫЙ УЧАСТОК (петля псевдоуридина) всегда содержит последовательность 5 '-TЦГ-3', этой петлей взаимодействует с рибосомой. АНТИКОДОНОВАЯ ПЕТЛЯ- содержит 3 нуклеотида, комплементарных кодону данной аминокислоты в мРНК, например, кодону 5'-ГЦЦ-3' в мРНК соответствует антикодон 3'-ЦГГ-5' в тРНК, чем обеспечивается специфичность взаимодействия с матричной РНК.
ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА ДНК
Молекулярная масса ДНК определяется рядом методов, в том числе:
а) Ультрацентрифугированием в градиенте CsCl (ММ от 200000 до 109);
б) по вязкости растворов (ММ> 109,
поскольку при центрифугировании такие длинные молекулы разрываются под действием
собственного веса).
ДЕНАТУРАЦИЯ ДНК заключается в разрыве Н-связей и стэкинг-взаимодействий, что приводит к расплетанию и разделению цепей. (БЕЗ РАЗРЫВА КОВАЛЕНТНЫХ СВЯЗЕЙ!) под действием температуры или рН.
О степени денатурации судят по
изменению интенсивности поглощения в ультрафиолете при l=
260 nm, поскольку дезэкранирование азотистых оснований в результате расплетания
цепей устраняется, что вызывает увеличение степени поглощения раствором ДНК
ультрафиолета указанной длины волны:
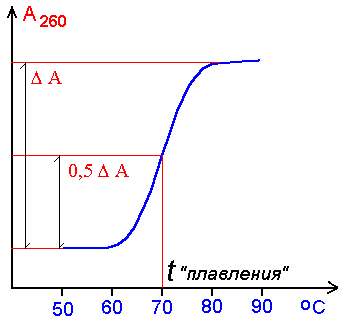
Денатурацию иначе называют ПЛАВЛЕНИЕМ, а температура плавления соответствует моменту 50%-ной денатурации молекулы. Т плав. разная для каждой ДНК.
Комплементарные цепи ДНК, разделенные при денатурации, при
определенных условиях могут вновь соединиться в двойную спираль. Этот процесс
называется ренатурацией. Если денатурация произошла не полностью и хотя бы несколько
оснований не утратили взаимодействия водородными связями, ренатурация протекает
очень быстро. Ренатурация возможна даже при полностью разделенных цепях. В таком
случае ренатурация требует точного совмещения цепей ДНК, которое может привести
к реассоциации, и этот процесс медленный, к тому же, зависит от концентрации
цепей в растворе. Как правило, выдерживание раствора ДНК при температуре на
10-15 С ниже температуры плавления в условиях средней ионной силы (0,15 М) обеспечивает
наиболее благоприятные условия для ренатурации. При более низкой ионной силе
ренатурации мешает взаимное отталкивание фосфатных групп. Реассоциация начинается
со взаимодействия коротких комплементарных последовательностей нуклеотидов,
время существования которых в ассоциированном состоянии может оказаться непродолжительным,
если соседние участки ДНК окажутся некомплементарными. Процесс повторяется снова
и снова, пока не ассоциируют "нужные" участки. Однако, как они провзаимодействуют,
двойная спираль ДНК восстанавливается очень быстро. Ренатурация- двухстадийный
процесс. На первом этапе должны встретиться два комплементарных участка. Вероятность
такой встречи описывается уравнением второго порядка, когда взаимодействуют
ДВЕ цепи:
dt/dc = -kC2,
где k- константа скорости реакции ассоциации.
Интегрирование этого уравнения дает:
C/C0 = 1/(1 + kC0t),
где С- концентрация однонитевой ДНК в момент времени t, выраженная в
молях на литр (моль/л). C0- концентрация однонитевой ДНК в момент времени
0. Построение зависимости 1-C/C0 (которая пропорциональна концентрации
реассоциированной ДНК) против значения C0t дает рисунок, показанный
ниже:

Доля наполовину реассоциированной ДНК определяет значения
C0t, соответствующего C/C0=0,5,
позволяет вычислить значение C0t0,5. Эта величина
пропорциональна сложности генома. СЛОЖНОСТЬ генома, в свою очередь, численно
равна его молекулярной массе (под термином "молекулярная масса" в случае ДНК чаще всего понимается число пар
оснований, которые она содержит), при условии, что этот геном единственный и имеет
УНИКАЛЬНУЮ последовательность. Например, сложность и молекулярная масса генома,
состоящего всего из трех ни разу не повторяющихся уникальных последовательностей нуклеотидов могут быть выражены
суммой нуклеотидных пар последовательностей: N1 + N2 + N3. В этом случае сложность и молекулярная масса генома совпадают. Однако, геномы эукариот,
содержащие как уникальные, так и повторяющиеся последовательности, имеют различные
сложность и молекулярную массу. При этом сложность их намного меньше молекулярной
массы. Например, если геном содержит 100000 копий последовательности N3,
1000 копий последовательности N2 и одну уникальную последовательность
N1, его сложность будет выражена суммой пар оснований последовательностей N1
+ N2 + N3, а молекулярная масса- суммой пар оснований 100000 N3
+ 1000 N2 + N1.
Суммарная "длина" генома сложноорганизованного человека и, например, лосося или комара, могут быть соизмеримы, однако, сложность генома человека намного выше. В геноме того же лосося, например, существует огромное число повторений одинаковых последовательностей нуклеотидов, вызванные физиологическими особенностями, средой обитания и т.д. Совершенно бессмысленно сравнение уровня развития организмов по числу хромосом или количеству генов (последовательностей нуклеотидов) в геноме, которые могут состоять из различного числа пар нуклеотидов.
C0t- кривые геномов эукариот с повторяющимися
фрагментами содержат несколько кинетических компонент. Наиболее часто повторяющиеся
последовательности реассоциируют быстрее всего:

Этот идеализированный график представляет ДНК, состоящую из
трех определенных компонентов последовательностей с тремя различными значениями
C0t. Их процентное соотношение можно прочитать на оси ординат
(1-С/Со). Частота повторения и сложность генома вычислены по принципам, описанным
выше:
| быстро реассоциирующие компоненты | средне реассоциирующие компоненты | медленно реассоциирующие компоненты | |
| Содержание в геноме, % | 25 | 30 | 45 |
| C0t0,5 | 0,0013 | 1,9 | 630 |
| Сложность, пар оснований | 340 | 6,0 х 105 | 3,0 x 108 |
| Частота повторения в геноме | 500 000 | 350 | 1 |
Зависимости для реальной молекулы ДНК совсем не так однозначны и легко читаемы,
как на показанном гипотетическом примере.
ОПРЕДЕЛЕНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ И НУКЛЕОТИДНОГО СОСТАВА
Наверх
Определенные сложности при определении нуклеотидной последовательности
вызваны тем, что имеется всего четыре кодирующих "буквы" и существует
высокая вероятность самопроизвольного разрыва молекул по сахарофосфатному остову,
даже в результате очень аккуратного пипетирования, из-за их огромных размеров.
Долгое время это сдерживало работы по определению полной нуклеотидной последовательности
человеческого генома.
Перед определением полинуклеотидной последовательности молекулы НК “разрезают” на более мелкие участки, которые затем разделяют, как правило, электрофорезом на полиакриламидном геле (ПААГ, PAGE, SDS-PAGE) или капиллярным электрофорезом (CE). При "резке" НК применяют, главным образом, особого рода ферменты, выделенные из различных микроорганизмов, которые называют "рестирикционными энзимами" или просто РЕСТРИКТАЗАМИ. Другое их наименование (более научное)- ЭНДОНУКЛЕАЗЫ. В настоящее время доступно большое число коммерческих рестриктаз. Название рестриктазы складывается из латинских букв и цифр, отражающих лишь их происхождение (буквы) и разновидность (цифры). Например, эндонуклеаза Pst I находит последовательность нуклеотидов CTGCAG на одной из цепей (на комплементарной цепи ей соответствует последовательность GACGTC) и разрезает каждую из цепей между нуклеотидами А и G. В результате получаются отрезки ДНК с "липкими" концами:

Название Pst I образуется из названия источника энзима- микроорганизма Providencia stuartii.
Существуют эндонуклеазы, разрезающие ДНК с образованием тупых концов, например:

вверх
процессинг нуклеиновых кислот
секвенирование ДНК
рекомбинантные ДНК и биотехнология
трансляция
процессинг ДНК в эукариотических
системах
вернуться на начальную страницу