К настоящему моменту времени установлено строение иммуноглобулинов класса G. Строение иммуноглобулинов других классов весьма совпадает со строением IgG. Все они имеют по 2 тяжелых и по 2 легких цепи, соединенных дисульфидными мостиками, как указано на схеме.

Средняя длина тяжелой цепи составляет 440 аминокислотных остатков (масса ~ 50 kDa), легкой- 220 (MM ~ 25 kDa).
Всего насчитывается пять классов иммуноглобулинов (IgA, IgG, IgD, IgE, IgM):
|
Класс Ig |
ММ, kDa |
Изотип и масса тяжелой цепи, kDa |
Сод-е углеводов, % |
Содержание в сыворотке, г/л |
Примечание |
|
G |
150 |
g, 53 |
2-3 |
6,0-18,0 |
|
|
A |
170-720 |
a, 64 |
7-12 |
0,9-4,2 |
[(LH)2]2 |
|
D |
160 |
d, 58 |
- |
0,003-0,4 |
|
|
E |
190 |
e, 75 |
10-12 |
0,0001-0,1 |
|
|
M |
950 |
m, 70 |
10-12 |
0,5-1,9 |
[(LH)2]5 |
В каждом классе иммуноглобулинов имеются подклассы (IgG1, IgG2, IgG3 …) и подподклассы (IgG1a, IgG2b и т. д.).
На молекуле антител различают КОНСТАНТНЫЕ (СН1, CH2, CH3, CL) и ВАРИАБЕЛЬНЫЕ (VH и VL) участки легкой и тяжелой цепей. Гипервариабельная зона вариабельного участка и является антигенсвязывающей (Fab)
. Молекула иммуноглобулина насчитывает, таким образом, ДВЕ зоны связывания антигена. При разрушении ферментом (папаин) молекулы антитела в зоне “шарнира”, образуется 3 фрагмента Ig- один константный Fc и два антигенсвязывающих Fab, сохраняющих свою способность связывать антигены.
ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ (ИФА, ELISA-Enzyme-Linked Immunoadsorbent Assay, EIA- Enzyme Immuno Assay)
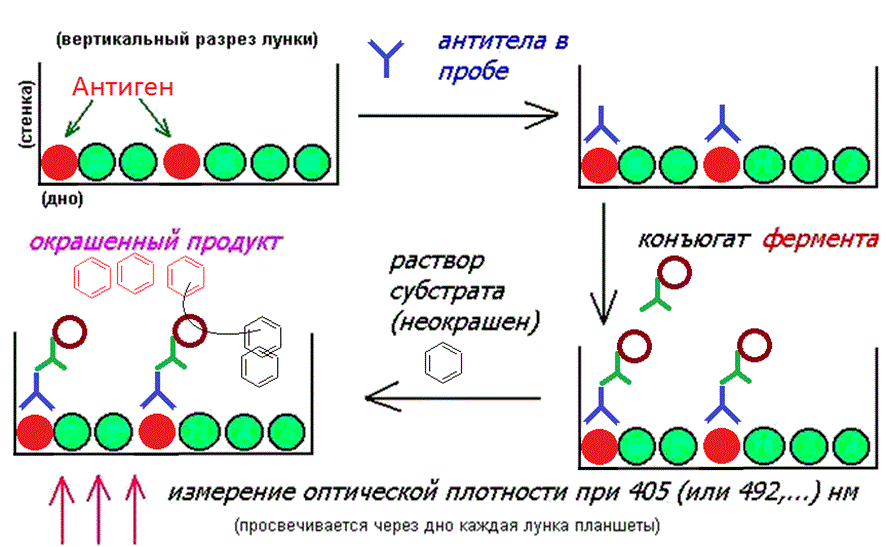
Применяется в диагностических целях, при определении в сыворотке крови специфических антител к возбудителю подозреваемой патологии. Используются специальные планшеты из полистирола, с круглыми плоскодонными лунками, вместимостью около 300 мкл. Число лунок на планшете- 96 (8 рядов по 12 лунок).
Перед анализом планшету сенсибилизируют, нанося раствор антигена. Обычно это не целая структура возбудителя, а часть (наиболее характерная) белков его оболочки. Очень часто антигены для ИФА получают генноинженерными технологиями, выращивая на трансгенных E.Coli.
Растворимые антигены закрепляются на поверхности планшеты рядом специфических взаимодействий непосредственно из раствора. Для снижения помех участки незанятой антигеном поверхности “укрывают” нанесением раствора бычьего сыровоточного альбумина. Несвязанный материал удаляют и планшету сушат. Сенсибилизированные планшеты можно хранить при –18 С не более 2-х недель.
При выполнении анализа лунки заполняют исследуемым материалом (сыворотка крови), нанося его в разные лунки с дробным разбавлением (1/2, 1/4, 1/8, 1/16, 1/32 и т.д.), после чего выдерживают около 30 мин для связывания. Если в образцах имеются антитела, комплементарные нанесенному антигену, они образуют с ним прочные комплексы. Образцы из лунок удаляются и несвязавшийся материал удаляется трехкратной
промывкой буфером.
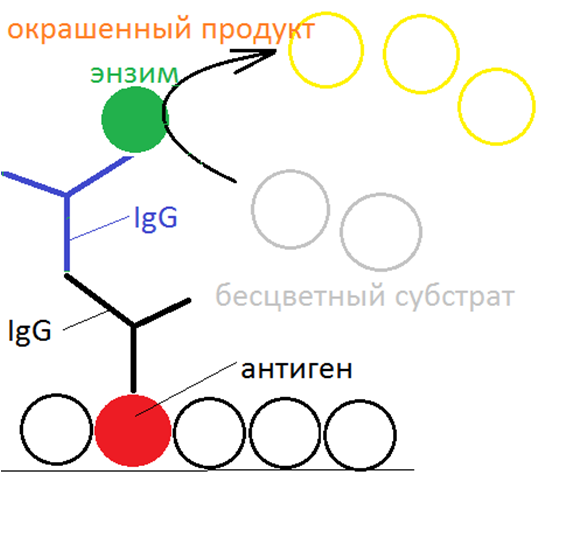
Далее лунки заполняются раствором конъюгата, представляющего собой связанные глутаровым альдегидом молекулы белка-фермента (чаще всего- пероксидаза хрена или щелочная фосфатаза) и молекулы антител против антител к возбудителю, полученные из сыворотки кролика (или другого организма) при введении в его кровь антител человека к искомому возбудителю. Дают время на прохождение реакции комплементарного связывания, после чего избыток раствора конъюгата
удаляется и лунки промываются буфером. Молекулы конъюгата осаждаются на молекулах осадившихся на антигене антител из испытуемого образца. В довершение всего лунки заполняют раствором субстрата, превращение которого в окрашенный продукт осуществляется конъюгированным ферментом. Такое ферментативное превращение осуществляется примерно 60 мин, после чего реакция останавливается добавлением кислоты. Количество превращенного субстрата пропорционально концентрации
конъюгата и, следовательно, концентрации антител против возбудителя в исследуемом образце. По плотности окраски в УФ-свете можно судить о концентрации антител в исследуемой сыворотке. В “холостых” контрольных лунках параллельно проводят реакцию для установления порога “шумов”. Имеются также лунки, заполненные антителами из контрольного раствора для проверки специфичности реакции.