| томский государственный университет |
| кафедра органической химии |
|
ОБМЕН БЕЛКОВ И АМИНОКИСЛОТ
|
За сутки в организме взрослого человека
обновляется до 400 г белка, с разной скоростью, причем 100 г из них распадается
необратимо. Эти 100 г белка (аминокислот) должны быть компенсированы.
Компенсация происходит частично за счет эндогенно
синтезируемых белков, частично- поступает с пищей. Пищевая ценность белоксодержащих
продуктов неоднородна.
|
Продукт |
содержание белка, г/100 г сырого продукта |
пищевая ценность |
|
мясо |
16-24 |
высокая |
|
рыба |
16-21 |
высокая |
|
сыры сычужные |
20-35 |
высокая |
|
яйцо |
11-14 |
очень высокая (эталон) |
|
молоко |
3,5 |
высокая |
|
картофель |
1,5-2 |
низкая |
|
соя |
32-37 |
низкая |
|
горох |
24-27 |
низкая |
|
гречневая крупа |
11-14 |
низкая |
|
хлеб ржаной |
7-8 |
низкая |
Пути поступления аминокислот:
а) транспорт из внеклеточной жидкости (при всасывании
пищевых аминокислот);
б) синтез заменимых аминокислот;
в) внутриклеточный гидролиз белков.
Пути потребления аминокислот:
а) синтез
белков и пептидов;
б) синтез небелковых азотсодержащих соединений (никотинамид, КоА, фолиевая кислота, адреналин, норадреналин, ацетилхолин);
в) синтез углеводов с использованием
углеродных скелетов аминокислот;
г) синтез липидов с использованием ацетильных остатков углеродных скелетов аминокислот;
д) окисление до конечных продуктов обмена.
Распад белков до аминокислот происходит при помощи ферментов ПРОТЕИНАЗ или КАТЕПСИНОВ.
ТРИ ГРУППЫ ПУТЕЙ РАСПАДА АМИНОКИСЛОТ
1. превращения NH2-
группы
2. распад углеродного скелета
3. декарбоксилирование
1. ПРЕВРАЩЕНИЯ АМИНОГРУППЫ
а) восстановительное дезаминирование
R-CH(NH2)-COOH + 2H ®
R-CH2-COOH + NH3
б) гидролитическое дезаминирование
R-CH(NH2)-COOH + HOH ®
R-CH(ОН)-COOH + NH3
в) внутримолекулярное дезаминирование
R-CH(NH2)-COOH ®
R-CH=CH-COOH + NH3
г) окислительное дезаминирование
R-CH(NH2)-COOH + 1/2O2
® R-C(=О)-COOH
+ NH3
Окислительное дезаминирование характерно
для большинства организмов и бывает 2-х видов:
а) прямое
б) непрямое (трансдезаминирование)
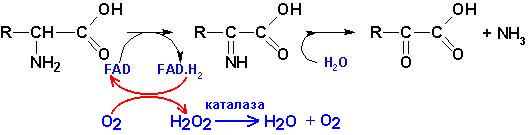
А) прямое окислительное
дезаминирование (оксидазами аминокислот), коферменты- FMN,
FAD:
Образующиеся при этом восстановительные эквиваленты FMN.H2
окисляются каталазой в присутствии кислорода воздуха, конечный продукт- перекись
водорода.
Б) трансдезаминирование (основной путь катаболизма),
протекает при участии аминотрансфераз или трансаминаз:
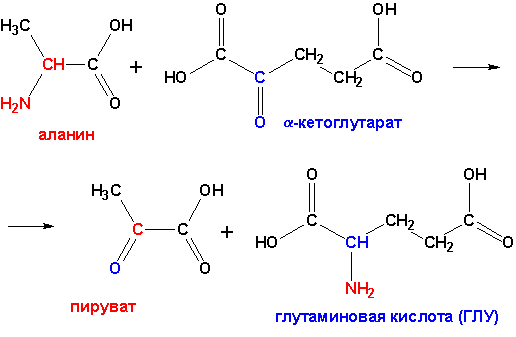
Аналогично происходит превращения с аспарагиновой кислотой:
Аспарагиновая
кислота (АСП) + a-кетоглутарат
® оксалоацетат
(ЩУК) + ГЛУ
Коферментами аминотрансфераз являются производные витамина В6 (пиридоксина)-
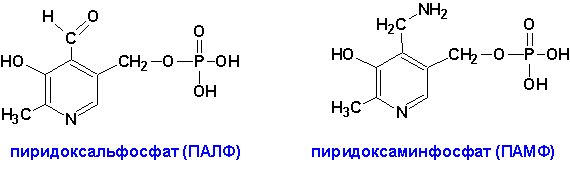
Для катализа требуются ОБА кофермента одновременно. В условиях организма они легко синтезируются из В6 и переходят друг в друга.
Механизм трансдезаминирования:
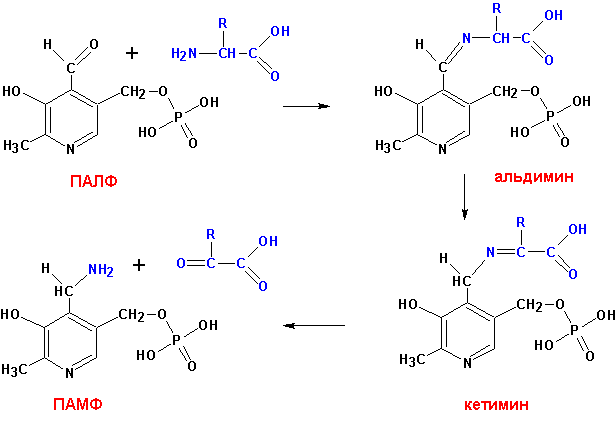
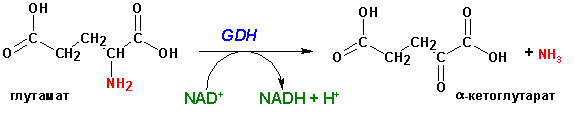
В условиях организма весь азот сосредотачивается
в глутаминовой кислоте, которая в митохондриях подвергается окислительному дезаминированию
при помощи NAD+, NADP+ - зависимой ГЛУТАМАТДЕГИДРОГЕНАЗЫ
(GDH):
МЕХАНИЗМЫ ОБЕЗВРЕЖИВАНИЯ АММИАКА (весьма
токсичного, особенно для нервных клеток, продукта):
а) восстановительное аминирование (трансреаминирование);
б) образование амидов аминокислот (АСН и ГЛН);
в) образование аммонийных солей;
г) образование мочевины (основной путь).
а) Восстановительное аминирование- малоэффективный процесс связывания аммиака, так как для этого необходимы значительные количества a-кетоглутарата.
б) Образование аспарагина (АСН) и глутамина (ГЛН) из соответствующих аминокислот АСП и ГЛУ является процессом получения амидов и протекает путем связывания аммиака при участии энергии АТФ.
в) образование аммонийных солей является процессом,
обратным “б”. АСН и ГЛН являются транспортными
формами аммиака, превращаясь в почках в результате гидролиза в соли аммония
и АСП и ГЛУ, соответственно.
Наиболее эффективным способом обезвреживания
аммиака в организме человека являются превращения с образованием мочевины. Мочевина
образуется из аммиака в результате функционирования орнитинового
цикла.
Превращения орнитинового цикла
протекают в цитоплазме, в то время как регенерируется аммиак в матриксе митохондрий.
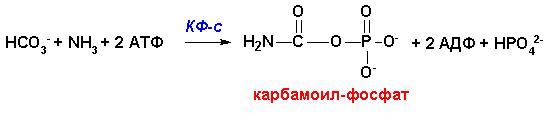
Орнитиновому циклу превращений предшествует реакция синтеза
карбамоилфосфата из аммиака и бикарбонат-иона, катализируемая ферментом
карбамоил-фосфат синтетазой
I(на
рисунке- КФ-с), которая не является
участником цикла мочевины (орнитинового цикла). Реакция с ее участием протекает
в матриксе митохондрий, аллостерическая регуляция- абсолютная и осуществляется
N-ацетилглутаминовой кислотой:
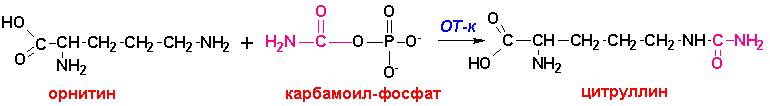
Аммиак пересекает внутреннюю митохондриальную мембрану (ВММ) в
составе аминокислоты цитруллин, в которой
он утилизируется при реакции карбамоил-фосфата с орнитином, поступающим из цитоплазмы.
Реакция катализируется ферментом орнитин-транскарбамоил-синтетазой
(ОТ-к):
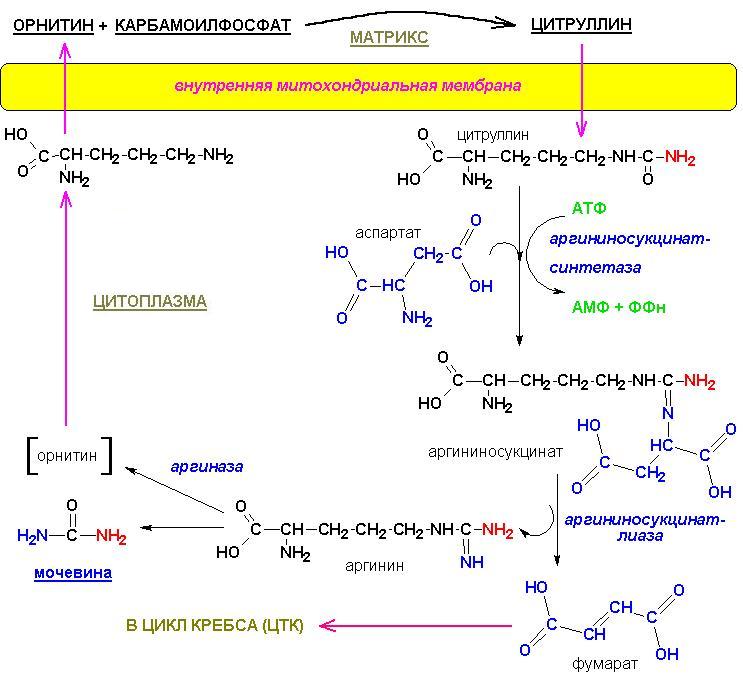
ОРНИТИНОВЫЙ
ЦИКЛ
Цитруллин пересекает митохондриальную мембрану и попадает в
цитоплазму, где протекает большая часть превращений орнитинового цикла:
ВЕРНУТЬСЯ НА НАЧАЛЬНУЮ
СТРАНИЦУ
|
© khassanov, MMIII-MMXVII
|