|
томский
государственный университет
|
|
кафедра
органической химии
|
|
МЕТАБОЛИЗМ
ЖИРНЫХ КИСЛОТ. ЦИКЛ КНООПА-ЛИНЕНА
|
Начинается с гидролиза ТРИАЦИЛГЛИЦЕРИНОВ под действием фермента
ТРИАЦИЛГЛИЦЕРИНЛИПАЗЫ. ФОСФОГЛИЦЕРИДЫ гидролизуются с помощью ФОСФОЛИПАЗ и локализованы в лизосомах.
СФИНГО- и ГЛИКОЛИПИДЫ- имеют свои ферменты.
СКОРОСТЬ ГИДРОЛИЗА СБАЛАНСИРОВАНА СО
СКОРОСТЬЮ ОКИСЛЕНИЯ.
Продукты гидролиза- ГЛИЦЕРИН и ЖИРНЫЕ КИСЛОТЫ- поступают из жировой ткани в кровь и далее к месту потребления.
ОБМЕН ГЛИЦЕРИНА тесно связан с гликолизом и дает 19 мол. АТФ в аэробных условиях.
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
(происходит только в митохондриях)
1) АКТИВИРОВАНИЕ ЖК
происходит в ЦИТОПЛАЗМЕ при помощи фермента ацил-КоА-синтетаза
с участием инонов Mg2+,
по схеме:
СН3(СН2)nСООН + АТФ + КоА-SHà
СН3(СН2)nСО-S-KoA + АМФ + ФФн
2) ТРАНСПОРТ внутрь митохондрий (с участием карнитина)
СН3(СН2)nСО-S-KoA
+ (СН3)3N+СН2СН(ОH)СН2СОO-
-->
(СН3)3N+СН2СН(ОCOR)СН2СОO- + KoA-SH
ЖК мигрирует через мембрану митохондрии с карнитином, после чего происходит обратный обмен с молекулой КоА-SH из матрикса.
В матриксе происходит окисление в цикле КНООПА-ЛИНЕНА.
СН3(СН2)n-СН2-СН2-СО-S-KoA+ FAD (ацил-КоА-дегидрогеназа)-->
СН3(СН2)n-СН=СН-СО-S-KoA+
FAD.Н2
¯ +
Н2О
(еноил-КоА-гидратаза)
СН3(СН2)n-СН(ОН)-СН2-СО-S-KoA
¯
+ NAD (-NADH) (3-гидроксиацил-КоА-дегидрогеназа)
СН3(СН2)n-СО-СН2-СО-S-KoA
¯
+ КоА-SH (ацетил-КоА-ацилтрансфераза)
СН3(СН2)n-СО-S-KoA + СН3СО-S-KoA
¯¯¯
СН3СН2СН2-CО-S-KoA
¯
2 СН3-CО-S-KoA
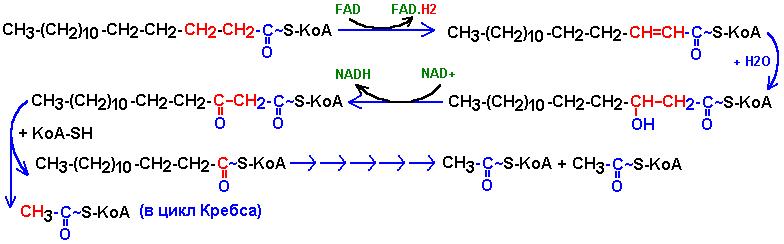
Продукты окисления FADH2 и NADH- в дыхательную цепь, ацетил-КоА- в цикл КРЕБСА.
ОСОБЕННОСТИ ОКИСЛЕНИЯ
а) ОКИСЛЕНИЕ С НЕЧЕТНЫМ ЧИСЛОМ С - образуется одна молекула ПРОПИОНИЛ-КоА, которая затем превращается в СУКЦИОНИЛ-КоА:
СН3СН2-CО-S-KoA
¯ +
СО2,
+ АТФ (-АДФ, -Фн, пропионил-КоА-карбоксилаза)
СН3СН(СООН)-CО-S-KoA
(метилмалонил-КоА)
¯ (метилмалонил-КоА-мутаза)
СООН-СН2СН2-CО-S-KoA
(сукциoнил-КоА---- в ЦТК)
б) окисление разветвленных жирных кислот
в) ОКИСЛЕНИЕ ЖК С НЕНАСЫЩЕННЫМИ
СВЯЗЯМИ- если двойная связь стоит там, где требуется на стадии енолизации,
то все идет как обычно, в случае ЖК с нас. связями. Если не в положении ТРАНС-
и не в том месте (D2,3),
то начинает работать фермент D
3,4-цис-D
2,3-транс-еноил-КоА-изомераза,
которая перемещает двойную связь и меняет ее конфигурацию.
Скорость окисления ненасыщенных
ЖК выше, чем у насыщенных:
СТЕАРИНОВАЯ (C18, все связи насыщенные)- относительная скорость-1
ОЛЕИНОВАЯ (С18, 1 двойная связь)- 11
ЛИНОЛЕВАЯ (С18, 2 двойные связи) - 114
ЛИНОЛЕНОВАЯ (С18, три двойные связи) - 170
АРАХИДОНОВАЯ (С20, четыре двойные связи) - 200
БАЛАНС ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ- на 1 молекулу ЖК с числом атомов С=n образуется 17(n/2)-6 молекул АТФ (пальмитиновая, n=16, дает 130 молекул АТФ. Энергетическая ценность ЖК (45 молекул АТФ на капроновую кислоту С=6) выше, чем у углеводов (38 мол АТФ на глюкозу). Однако, при сгорании в ЦТК молекул ацетил-КоА требуется достаточное количество ОКСАЛОАЦЕТАТА (ЩУК), источником которых являются УГЛЕВОДЫ. Таким образом ЖИРЫ СГОРАЮТ В ПЛАМЕНИ УГЛЕВОДОВ.
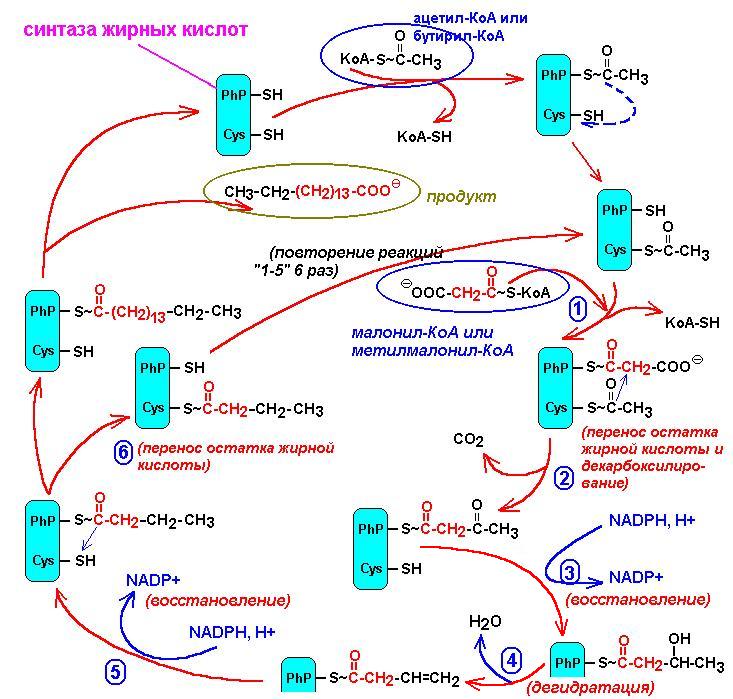
БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ
Легко протекает в любом организме (в цитоплазме). Материал- ацетил-КоА.
Условия- обильное потребление углеводов, накопление изоцитрата, цитрата, фумарата.
СН3-CО-S-KoA + НСО3- + АТФ
¯ карбоксилаза
НООС-СН2-CО-S-KoA + АДФ + Фн (малонил-КоА)
¯ (+СН3-CО-S-KoA,
+ АПБ-(жирнокислотная синтаза), простетическая
группа-4-фосфопантотеновая кислота (PhP)
и тиоэтиламин (Cys)
НООС*-СН2-CО-S-АПБ + СН3-CО-S-АПБ
¯
СН3-CО-СН2-CО-S-АПБ
+ С*O2 (ацетоацетил-S-АПБ)
¯ +
NADPH, - NADP
СН3-CH(ОH)-СН2-CО-S-АПБ
(3-гидроксибутирил-АПБ)
¯ -Н2О
СН3-CH=СН-CО-S-АПБ
(кротонил-АПБ)
¯ + NADPH, - NADP
СН3-CH2-СН2-CО-S-АПБ
(бутирил-АПБ)
Таким образом циклы повторяются
до тех пор, пока не будет синтезирована цепь в 16 атомов углерода.
На рисунке приведена схема синтеза жирных кислот.
БИОЭНЕРГЕТИКА
ГЛИКОЛИЗ
ЦИКЛ ТРИКАРБОНОВЫХ
КИСЛОТ
МЕТАБОЛИЗМ
ЧЕЛНОЧНЫЕ МЕХАНИЗМЫ ТРАНСМЕМБРАННОГО
ПЕРЕНОСА NADH